4. Růst mikroorganizmů Microbial growth
Scene 1 (0s)
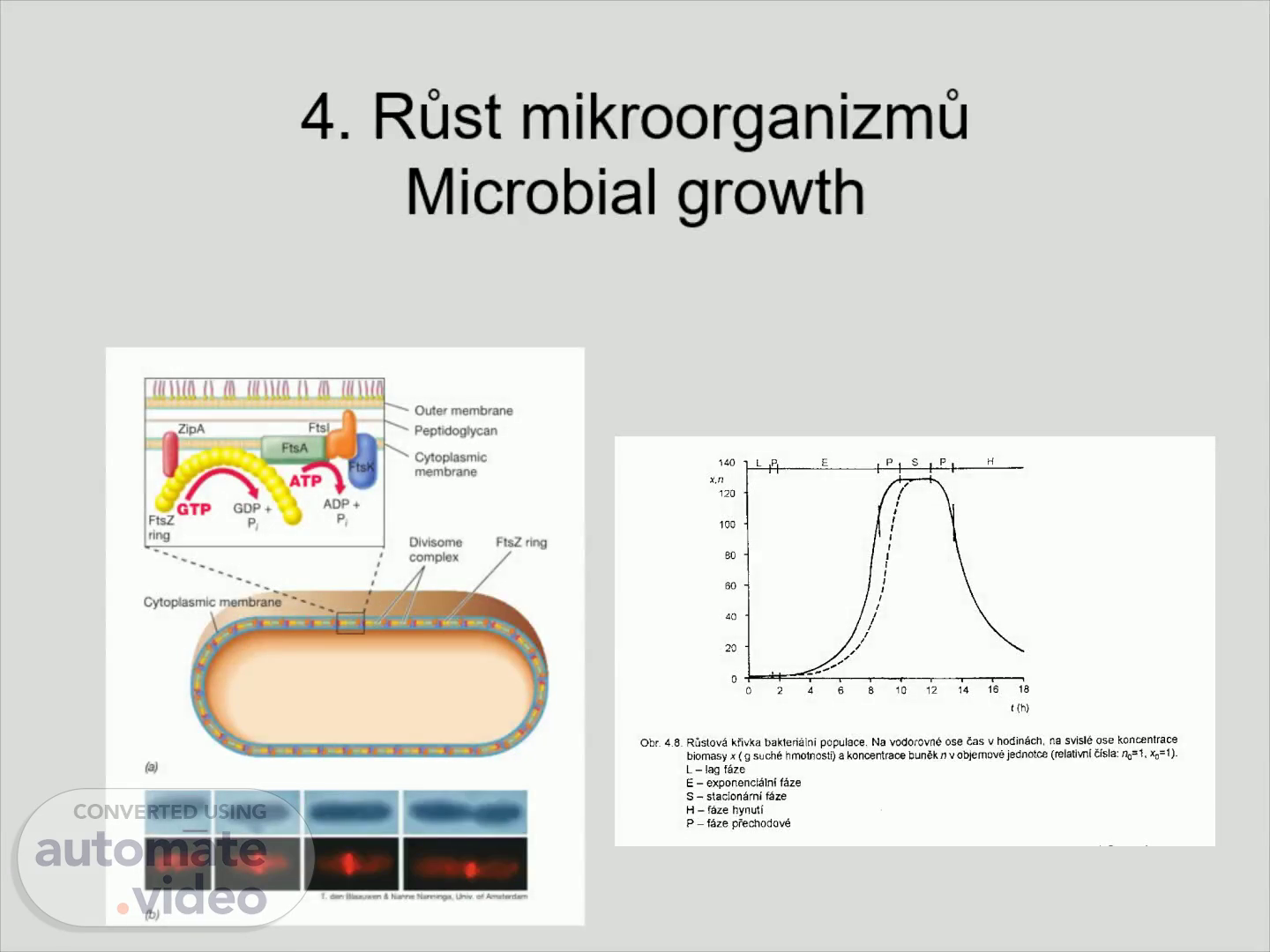
4. Růst mikroorganizmů Microbial growth. 4. GTP Cyto mem b rane ' • ATP GDP co m plex FtSZ.
Scene 2 (58s)
Schéma přednášky. Buněčné dělení u bakterií Růst populace mikroorganizmů Měření růstu populací mikroorganizmů Teplota a růst mikroorganizmů Ostatní faktory prostředí a růst mikroorganizmů Inhibice růstu mikroorganizmů.
Scene 3 (2m 5s)
**Literatura. Microbes and evolution (2012)_ Lenski experiment and salute to Darwin Encyclopedia of Microbiology (3rd ed.)_Chemical agents.
Scene 4 (3m 18s)
Růst buňky mikroorganizmu a růst populace. Výchozí stav - individuální růst: Model: bakterie – Escherichia coli Buňka v ideálním médiu Růst a dělení buňky Charakter dělení Individuální růst velmi dobře prozkoumaný na molekulární úrovni F.Jacob : Po čem by měla toužit bakteriální buňka než po dělení na dvě buňky? -- Populace mikroorganizmů – obecné rysy: Vznik populace, vývoj populace v čase Matematické modely růstu – nejlepší popis pro praktické použití.
Scene 5 (6m 26s)
Dělení tyčinkovité bakterie Etapy individuálního růstu.
Scene 6 (8m 47s)
Etapy dělení bakteriální buňky. Klíčové procesy v buněčném cyklu: Replikace DNA a rozdělení genetického materiálu Tvorba septa (přepážky) Oddělení buněk Tyto procesy musejí proběhnout v definovaném pořadí a musejí být citlivě regulovány Nesmyslná strategie: Tvorba septa a oddělení před dokončením replikace.
Scene 7 (10m 17s)
Buněčný cyklus u Escherichia coli .. Regulace - základní momenty: Replikace se zahajuje po dosažení tzv. kritického objemu buňky Po dokončení replikace se zahajuje tvorba přepážky Dvě fáze buněčného cyklu: fáze C: syntéza DNA, fáze D: tvorba přepážky + oddělení buněk C: 41 min. D: 20 min. C +D ~ 60 min. (Ještě se rozeznává „Fáze B“: od oddělení buněk k počátku replikace, existuje u pomaleji se dělících při T <60 min) Generační doba T může být < C+D (např. 40 min., minimálně 20 min.) TEHDY PLATÍ: Zahájení replikace : každých T minut C a D fáze se mohou překrývat (replikace během tvorby septa) Syntéza DNA „dcery“ se zahajuje v „babičce“.
Scene 8 (15m 8s)
Replikace bakteriálního chromozomu. Genom bakterie: 1 cyklický bakteriální chromozom (nukleoid) Replikace: jediný origin (replikační počátek) Replikace běží oběma směry („ theta struktura“) Každý nukleoid je během replikace připojen k CPM a zreplikovaná DNA putuje do jednoho z pólů buňky.
Scene 9 (17m 30s)
Cytoskeletální proteiny u bakterií. Bakteriální buňka není „balík enzymů“, ale buňka s orientovanými strukturami – póly, střed buňky, tvorba přepážky, distribuce chromozomů Za orientaci jsou zodpovědné cytoskeletální proteiny Všechny cytoskeletální proteiny eukaryot mají bakteriální homology Homology plní jinou funkci než u eukaryot Obecná funkce těchto proteinů: tvorba dynamických vláknitých struktur.
Scene 10 (20m 29s)
Tvorba přepážky a proteiny Fts. Fts = F ilamentous T emperature S ensitive Proteiny Fts tvoří společně DIVISOM FtsZ : Příbuznost s tubulinem , GTPáza Výskyt: FtsZ u Bacteria i Archaea , ostatní Fts pouze u Bacteria.
Scene 11 (22m 3s)
Schéma divizomu Escherichia coli. Prstenec z FtsZ ve středu buňky cca 10 000 molekul FtsZ ZipA , FtsA – spojení CPM a FtsZ FtsA – homolog aktinu FtsI – syntéza peptidoglykanu , jeden z PBP proteinů ( P enicillin - B inding - P rotein) FtsK – táhne nukleoidy k pólům buňky (účastní se i Mre a Par proteiny).
Scene 12 (25m 19s)
Návaznost replikace a tvorby přepážky. Prstenec FtsZ tvořen tam, kde není nukleoid (póly a střed u téměř zreplikované DNA) a tam, kde nejsou proteiny Min Oscilace prstenců MinC , MinD a MinE mezi oběma polovinami buňky Výsledek: Prstenec FtsZ tvořen ve středu buňky.
Scene 13 (28m 2s)
Proteiny Min a tvorba prstence FtsZ .. Prstenec FtsZ je tvořen tam, kde není nukleoid (póly, střed) MinC inhibuje polymeraci FtsZ . Proteiny MinC a MinD vytvářejí na pólu strukturu na cytoplazmatické membráně, která je degradována MinE a která osciluje mezi oběma polovinami buňky. FtsZ tedy může vytvořit prstenec pouze ve středu buňky, kde je průměrná koncentrace MinC nejnižší..
Scene 14 (29m 55s)
Oscilace struktur tvořených MinC a MinD v buňce..
Scene 15 (31m 42s)
Tvar buňky a protein MreB Homologie s aktinem. MreB – spirála na vnitřní straně CPM, iniciace tvorby pepidoglykanu v místě styku s CPM MreB příbuzný aktinu , účast v distribuci nukleoidů k pólu (spolu s Par proteiny) Aktin – tvorba mikrofilament u eukaryot Tvorba bakteriální tyčinky vyžaduje MreB Koky nemají MreB , kulovitý tvar je „původnější“ – tyčinky mají kulovitý tvar při nefunkčním MreB.
Scene 16 (34m 10s)
Protein MreB a morfologie bakteriálních buněk. Tyčinka jako tvar bakterie – zodpovědný protein MreB Rotace spirály MreB , iniciace tvorby peptidoglykanu Asymetrie tyčinky (zahnutí buňky) – crescentin , Caulobacter Podobné proteiny u dalších zahnutých bakterií.
Scene 17 (36m 58s)
Dělení buňky a cytoskelet - Bacteria , Archaea , Eukarya.
Scene 18 (38m 12s)
Homologie eukaryotních a bakteriálních cytoskeletálních proteinů názorně.
Scene 19 (39m 26s)
Syntéza peptidoglykanu : tyčinky a grampozitivní koky.
Scene 20 (40m 53s)
Syntéza peptidoglykanu : přenos monomerů přes cytoplazmatickou membránu.
Scene 21 (43m 0s)
Syntéza peptidoglykanu : mechanizmus. Autolyziny – hydrolýza beta-1,4, vazby, hydrolýza spojení mezi oligopeptidy Transglykosylace – tvorba beta-1,4, vazby Transpeptidace – spojení sousedících oligopeptidů peptidickou vazbou (víc enzymů) Proteiny PBP – „ Penicillin binding proteins “ – vazba penicilinu na PBP inhibuje transpeptidační aktivitu Účinek penicilinu vyžaduje rostoucí buněčnou stěnu.